

Abstract
Sustainable agricultural production requires an inclusive framework that concurrently considers the impacts of production methods on soil health, crop productivity, and crop nutritional quality. However, few studies have directly examined the potential associations among management impacts on soil health (SH), crop productivity, and crop quality. We evaluated these linkages in a continuous maize experiment with treatments varying in nitrogen (N) fertilizer amount and type (zero input control, inorganic N, and manure-based N treatment). We evaluated select SH indicators after six cropping years, and computed physical, chemical, biological, nutrient, and overall SH indices using the Soil Management Assessment Framework (SMAF). Crop yields, mineral nutrient concentrations (denoted with brackets), and nutrient uptake were analyzed in years six and seven. Manure application increased biological SH indicators compared to the control and inorganic N treatments and also increased available potassium (K), zinc (Zn), copper (Cu), and phosphorus (P). Overall SH indices were higher in the inorganic N and manure treatments than in the control but did not differ between the two N sources, despite the large exogenous sources of C, N, and nutrients applied via manure. The SMAF tool only directly considers P and K in terms of soil nutrients—other nutrient benefits of manure application (i.e., increases in total soil N, available Cu and Zn) were not accounted for, suggesting that the SMAF nutrient SH index should be modified to account for the impacts of management practices on nutrient availability. Crop yields were higher in treatments with higher overall SH, supporting the linkages between SH and crop productivity. Despite widely differing nutrient inputs and soil fertility levels, we found no yield differences between the two N sources. However, there were notable treatment impacts on crop quality. Grain [N] was 40% greater in the urea and manure treatments than in the control. Grain [P], [K], and [Mg], important elements in livestock nutrition, were 10% to 28% greater in the manure treatment than in the urea treatment. In addition, although a yield dilution of grain [Zn] occurred in the urea treatment, the dilution effect was mitigated in the manure treatment, likely due to increases in available Zn. Overarchingly, our results suggest that management practices that maintain or improve SH and nutrient availability also improve maize productivity and nutritional quality, which could have cascading positive impacts on animal and human nutrition.
Introduction
Agriculture is, in the most basic sense, the provider of the essential nutrition for life. While extensive agricultural investment in the latter half of the twentieth century focused on increasing crop yields (Fuglie et al. 2012), production goals are expanding to include the additional targets of minimizing the environmental impacts of production, maintaining and improving soil health (SH), and sustaining or improving crop nutritional quality for human and animal health (Graham et al. 2001).
Soil health, also referred to as soil quality, is defined as “the continued capacity of soil to function as a vital living ecosystem that sustains plants, animals, and humans” (USDA NRCS 2019). Decades of research have focused on developing indicators and frameworks to evaluate SH (Bünemann et al. 2018; Doran and Parkin 1994; Moebius-Clune et al. 2016). Tools such as the Soil Management Assessment Framework (SMAF) integrate individual SH indicators into physical, chemical, biological, and nutrient index scores, which can be combined to produce an overall SH index (Andrews et al. 2004).
Numerous researchers have suggested that improvements in SH also lead to increases in crop productivity and nutritional quality (Pepper 2013; Wall et al. 2015; Warkentin 1995). Although improvements in SH can potentially promote crop yields in some systems via increased nutrient cycling and water capture, these linkages are only beginning to be quantitatively explored (Roper et al. 2017). However, increases in crop yields do not necessarily lead to increases in nutritional quality. The mineral nutritional component of crop quality (e.g., oil, protein, starch, or mineral macro- and micronutrients) is an enduring point of concern and interest, as deficiencies in nutrients, such as iron (Fe) and zinc (Zn), impact an estimated third to half of the global population (Miller and Welch 2013). Sufficient crop nutrient concentrations (denoted with brackets) are also necessary for meeting the nutritional needs of livestock (Gupta et al. 2008).
Maize (Zea mays L.) has been called a “paramount staple crop” in global nutrition, serving as a staple in the diet of over 200 million people, and provides an important entry point for nutrients into the global food chain (Nuss and Tanumihardjo 2010). Maintaining or increasing mineral nutrients in staple crops such as maize is a critical component of combating nutrient deficiencies, yet the interrelationships among management practices, SH, crop productivity, and crop quality are poorly understood.
Soil organic carbon (SOC) is widely considered the most important baseline measurement of SH due to its influence on multiple biological, chemical, and physical soil properties (Doran and Parkin 1994). Hence, when looking for linkages between SH, crop productivity, and crop quality, a good model system for study is one with treatments receiving exogenous sources of organic C (e.g., manure, other organic amendments) versus systems receiving only inorganic fertilizer or no additional inputs. Manure additions result in large direct C inputs that can increase SOC more effectively than mineral fertilizers (Maillard and Angers 2014; van der Bom et al. 2019). Increases in C via manure application can increase soil microbial biomass and activity (Kallenbach and Grandy 2011), with subsequent positive impacts on soil physical properties such as macro aggregation (Chaney and Swift 1984; Mikha and Rice 2004). In addition, manure contains other macro- and micronutrients, and repeated years of application effectively builds soil nutrient pools (Eghball and Power 1999b; Eghball et al. 2002).
However, there are tradeoffs between inorganic and manure-based N fertilization (Ribaudo et al. 2011). Notably, the nitrogen (N) in manure must be mineralized before it is plant available, making it difficult to ensure that N is available at critical crop growth stages (Eghball et al. 2002). Manure application rates that are sufficient to meet crop N needs can result in overapplication of phosphorus (P), as the crop N:P uptake ratio is higher than the ratio in manure (Schröder 2005). Elevated soil P concentrations are of environmental concern if P is moved via erosion into water bodies, where it can cause eutrophication (Hansen et al. 2002).
Real challenges and potential tradeoffs exist in production agriculture, underscoring the importance of concurrently assessing management impacts on SH, crop productivity, and crop quality. Our study objectives were (1) to evaluate changes in SH indicators and composite indices in response to soil amendments (zero N control, inorganic N treatment, and manure-based N treatment), and (2) to assess whether shifts in SH resulted in concurrent impacts on crop productivity and crop quality.
Materials and Methods
The study was located on a Fort Collins clay loam soil (fine-loamy, mixed, mesic Aridic Haplustalfs; 1% to 2% slope) at the Agricultural Research Development and Education Center near Fort Collins, Colorado. The study was initiated in 2012, with experimental plots laid out as a randomized complete block design with four replicates and multiple N source treatments (Halvorson et al. 2016). This experiment simulated a modified continuous corn production system, wherein stover was cut and baled after grain harvest, leaving only 5 to 7 cm stalk stubs in the field. Baseline soil properties in the 0 to 7.5 cm soil depth for the plot area were the following: pH at 8.0 (1:1); SOC at 11.9 g kg−1; total soil N at 1.5 g N kg−1; electrical conductivity (EC) at 0.58 dS m−1; bulk density at 1.34 g cm−3; sand at 409 g kg−1; and clay at 337 g kg−1. For the current study, we examine three N source treatments—a control treatment with no N applied, an inorganic N treatment (179 kg N ha−1), and a dairy manure treatment (target seasonal N availability of 179 kg N ha−1).
Each spring, manure was sourced from a local dairy and stockpiled in 2 × 20 m piles for ~30 days prior to application. The manure was mixed one to two times in that period to promote homogenization. One week prior to application, samples were taken to determine N content to calculate application rates. Due to the short mixing time and low air temperatures, we assumed that 40% of total N applied would be available during the first growing season, similar to estimates for fresh manure (Eghball and Power 1999b). The water content of the stockpiled manure was measured ~24 hours prior to application. Manure was hand applied to plots between March 15 and April 15. Four subsamples of the manure applied to each plot were also collected the day of application and composited. Samples were dried at 60°C until a constant weight, ground to pass a 150 um screen, and analyzed via combustion for total N and C (Elementar Americas Inc., Mt. Laurel, New Jersey). Samples from each plot were also sent to the Soil, Water, and Plant Testing Laboratory at Colorado State University for pH, total P, total potassium (K), extractable bases, total and available micronutrients, and soluble salt determination. Manure application rates, estimates of annual nutrient additions, and manure characteristics are presented in tables 1 and 2.
Manure was incorporated via rototiller the same day as application to a depth of ~15 cm (simulating tillage) to reduce N losses (Leikam and Lamond 2003). All plots in the study, including the urea and control treatments, were tilled the same day. Maize was planted into the tilled seedbed each year. In 2017, the hybrid was Channel 192-09VT3PRIB. This variety was unavailable for the 2018 growing season, so a variety was chosen that closely matched the 2017 hybrid (Channel 193-53STXRIB). Urea was broadcast at emergence each year at a rate of 179 kg N ha−1, with irrigation applied within two days. Triple superphosphate (0-46-0) was applied via surface broadcast to the urea and control treatments at a rate of 56 kg P ha−1 in 2013, 2015, and 2018 to avoid P deficiencies. Soil moisture was monitored with Watermark sensors (Spectrum Technologies Inc.), and water was applied as needed via a linear-move irrigation system. Growing season precipitation (April to October) totaled 298 mm in 2017 versus 192 mm in 2018. Total growing season water (precipitation + irrigation) totaled 673 mm in 2017 and 719 mm in 2018.
Soil Analyses. Soil samples were collected in May of 2018 after manure and urea applications. Two soil cores were collected in each plot (0 to 30 cm); separated into increments of 0 to 7.5, 7.5 to 15, and 15 to 30 cm; and composited. Samples were air-dried and then passed through an 8 mm screen, with a 50 g subsample taken for water-stable aggregate analysis using a modified Yoder sieving machine with nested sieves of 2, 1, 0.5, and 0.25 mm screen sizes (Kemper and Rosenau 1986). The remainder of each sample was passed through a 2 mm screen. Soil pH and EC were measured on a 1:1 soil to water basis. Sodium bicarbonate (NaHCO3) extractable soil P (Olsen et al. 1954) was analyzed via continuous flow analyzer (Lachat Instruments, Loveland, Colorado). Although inorganic N levels and available micronutrients are not directly included in the SMAF framework, these values were of interest to this study due to their impact on plant growth and crop productivity. Soil NO3 and NH4 were measured via continuous flow analyzer after extraction with 2 M KCl. Available soil Zn, manganese (Mn), Fe, copper (Cu), K, and magnesium (Mg) were analyzed via inductively coupled plasma optical emission spectrometry (ICP-OES) after diethylenetriaminepentaacetic acid (DTPA) extraction (Lindsay and Norvell 1978). Samples for total soil C (TSC) and total soil N (TSN) analyses were further ground to pass a 150 um screen before analysis (Elementar Americas Inc., Mt. Laurel, New Jersey). Soil inorganic carbon (SIC) was determined via pressure calcimeter (Sherrod et al. 2002). Soil organic carbon was calculated as the difference between TSC and SIC. β-Glucosidase (BG) activity was assayed using the methods of Eivazi and Tabatabai (1988). Potentially mineralizable N (PMN) was measured via a 28 day incubation—net mineralized N was calculated by subtracting the baseline mineral N from mineral N values obtained after incubation (Drinkwater et al. 1996). Bulk density (BD) was determined via the core method on one sample taken within each replicated plot after harvest in fall of 2018.
Annual manure application rates (dry weight basis) and estimated total and extractable (i.e., available) nutrients applied with manure (dry weight basis). No manure was applied in 2016.
Chemical and nutrient characteristics of manure by year. Only one composite sample was analyzed in 2012, except for carbon (C) and nitrogen (N). Values for all other years represent the average values from four sample analyses, and numbers in parentheses represent the standard deviation. Each of the four samples was aggregated from four subsamples taken from the manure applied to each field plot replicate. No manure was applied in 2016.
Soil Management Assessment Framework. We utilized the SMAF tool to calculate physical, chemical, biological, nutrient, and overall SH indices for each treatment by soil depth. Unitless scores for each SH indicator range from 0 to 1, with higher scores indicating “better” SH. Scoring curves are dependent on soil taxonomy and texture, temperature and rainfall regimes, slope, and crop. Scores for the indicators in each index are added together and divided by the numbers of indicators used to calculate that index (e.g., for the biological SH index, the indicator values for SOC (%), PMN, and BG are added together and divided by three). The overall SH index is calculated by summing the scores for each SH index category, multiplying by 100, and dividing by the total number of indicators. Full details of the SMAF algorithms are provided in Andrews et al. (2004) and Stott et al. (2010).
Plant Analyses. Aboveground biomass sampling occurred each year in late September at physiological maturity, prior to canopy senescence. Fifteen plants were harvested from a 1.5 m2 area within each plot. Plants were separated into grain (i.e., biomass grain), cobs, and stover (i.e., stalks plus leaves) and dried at 60°C for >72 hours for dry mass determination. Harvest grain yields were determined in late October by hand harvesting ears from an established plot area and separating the grain from the cob with a mechanical sheller. Final stover, cob, and harvest grain yields are expressed on an oven dry basis. Samples were dried and ground to pass a 150 um stainless steel screen and analyzed for C and N (Elementar Americas Inc., Mt. Laurel, New Jersey). Total macro- and micronutrients in each plant fraction were determined via ICP-OES after nitric acid (HNO3)–hydrogen peroxide (H2O2) digestion (Huang and Schulte 1985). The use of brackets [] around a nutrient denotes concentration. Each run included replicates and blanks, as well as replicates of certified plant standards (US National Institute of Standards). Total nutrient uptake in each plant compartment was calculated by multiplying biomass yields by nutrient concentrations.
Statistical Analysis. Soil data was analyzed separately for each depth using PROC MIXED in SAS v.9.4 (SAS Institute 2013) to test the significance of amendment on SH indicators and SMAF indicator scores. Amendment was considered a fixed effect, and block replication considered random. For the plant and yield data, repeated measures analysis of variance (ANOVA) was used in PROC MIXED to analyze treatment effects over years. It is important to note that while the hybrids utilized in 2017 and 2018 were closely matched in traits, target environment, and relative maturity, with presumably very similar genetics given that they came from the same breeding program, we cannot rule out the possibility of genetic differences contributing to differences in nutrient concentrations between growing years (Grusak and DellaPenna 1999). While differences between growing years are discussed when significant, directional trends in nutrient concentrations were consistent between years unless otherwise noted, and results are averaged over the two site years. Statistical comparisons for all data were made at the α = 0.05 probability level using the LSD method for means separation.
Results and Discussion
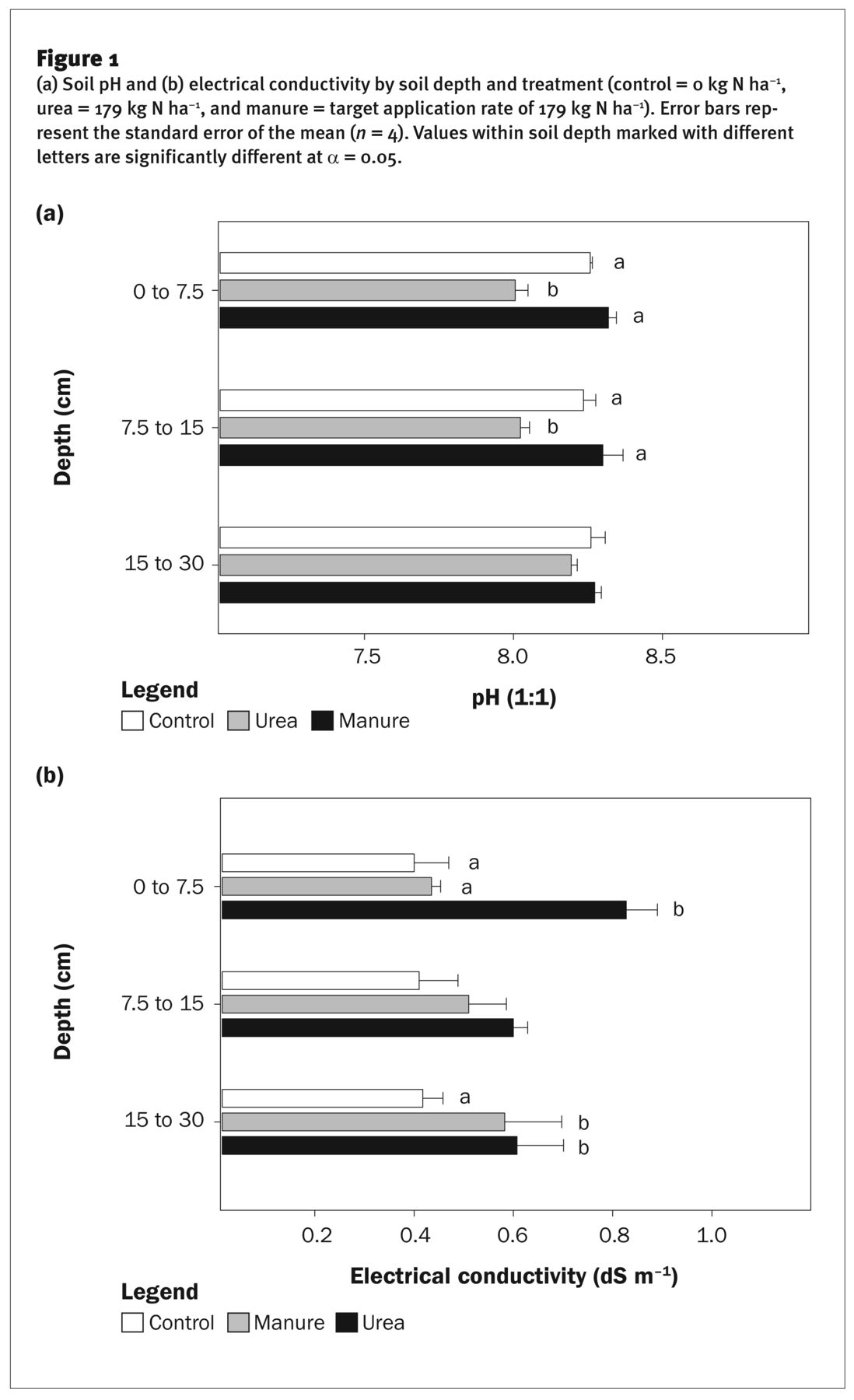
Soil Quality Indicators. Soil pH in the 0 to 7.5 cm and 7.5 to 15 cm depths of the control and manure treatments were similar, averaging 8.3, whereas pH in the urea treatment was 0.3 units lower (figure 1a). Surface soil pH in the urea treatment likely declined due to acidity produced following urea application (Havlin et al. 2005).
Soil EC in the surface 0 to 7.5 cm was 0.4 dS m−1 higher in the urea treatment than in the control and manure treatments (figure 1b). Soil EC measures the total concentration of soluble salts in solution, and the increase in EC in the urea treatment is likely due to higher soil nitrate (NO3−) (figure 3a). Manure can contain large amounts of salts, which can in some cases increase EC and negatively impact plant growth via water and ion imbalances. However, multiple studies have reported only nominal changes in soil EC after repeated years of manure application (i.e., <0.5 dS m−1) (Calderon et al. 2018; Eghball et al. 2004; Schlegel et al. 2017), and our results similarly indicate that manure application does not necessarily negatively impact soil EC. In addition, the general EC thresholds wherein maize yields are impacted are well above the EC levels we measured. Previously published salinity tolerance indices for maize developed under greenhouse conditions with chloride salts indicated initial yield declines at ≥1.70 dS m−1 (Maas and Hoffman 1977). However, recent work in field maize utilizing natural gradients of salinity found no declines in growth at EC values >9 dS m−1 (Butcher et al. 2018), indicating that tolerance thresholds in the SMAF framework may need to be reevaluated or calibrated for additional soils.
Manure amendment increased SOC (%) and TSN (%) in the surface 0 to 7.5 cm and 7.5 to 15 cm depths compared to the control and urea treatments, with no differences in the 15 to 30 cm depth (figure 2a). Due to the near-total aboveground biomass removal from silage harvest, the majority of C returned to the soil is root derived or via manure. Halvorson et al. (2016) reported increases in SOC and TSN in this system after three years of manure application, whereas SOC and TSN decreased or remained constant in the urea and control treatments. Manure contains large amounts of C and N (table 1), and increases in SOC and TSN following manure addition are well documented (Maillard and Angers 2014), underscoring the role manure can play in restoring or building SOC and TSN and overall SH.
Manure amendment increased BG activity by up to 50% in the 0 to 7.5 cm and 7.5 to 15 cm depth (figure 2c). In the 7.5 to 15 cm depth, BG activity of the urea treatment was higher than the control, likely due to increased root-derived C inputs. No treatment differences were detected in the 15 to 30 cm depth. β-glucosidase activity is a sensitive indicator of soil metabolic activity and C cycling—our results agree with other studies that have found that manure amendments increase BG activity (Acosta-Martinez et al. 2011; Calderon et al. 2018; Garcıa-Gil et al. 2000).
Potentially mineralizable N was higher in the urea and manure treatments than in the control in all three soil depths (figure 2d). Immobilization of N was evident in the control treatment (i.e., the conversion of inorganic N to organic N) for all three soil depths, reflecting the low inorganic N levels after seven years of no N fertilization.
There were no treatment differences in water-stable aggregates >250 μm at any depth (data not shown). While some studies have found increases in macroaggregates with manure application, these studies are often of longer duration. For example, Aoyama et al. (1999) and Mikha et al. (2015) reported increases in macro-aggregation after 18 and 70 years of manure application, respectively. Irrigated semiarid systems also appear to have relatively low levels of macroaggregation (Gillabel et al. 2007). In addition, tillage can disrupt or prevent the formation of stable macroaggregates (Mikha and Rice 2004; Six et al. 2000), and annual tillage events in this system may have minimized the positive impacts of manure amendments.

(a) Soil pH and (b) electrical conductivity by soil depth and treatment (control = 0 kg N ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 4). Values within soil depth marked with different letters are significantly different at α = 0.05.
Despite the large input of lower density organic matter (OM) via manure application, there were no treatment differences in BD at any depth (data not shown). These findings concur with those of Eghball (2002), who also reported no changes in BD with multiple manure applications in a similar system characterized by high clay content and low OM.
Soil Macronutrients. The bulk of inorganic soil N was in the form of nitrate nitrogen (NO3-N) (figure 3a); ammoniacal nitrogen (NH4-N) was <8 mg kg−1 in all depths and did not vary between treatments (data not shown). NO3-N was markedly higher in the urea treatment in all depths, ranging from 88 mg kg−1 in the surface 0 to 7.5 cm to 39.9 mg kg−1 in the 15 to 30 cm depth. Sampling occurred shortly after urea was applied, and hence these values reflect the high available N at emergence in this treatment. NO3-N levels were not different between the control and manure treatments, ranging from 7.2 to 19.8 mg kg−1, levels considered limiting to maize production (Davis et al. 2009).
Soil test P (STP) levels did not differ between the control and urea treatments at any soil depth; STP in the 0 to 7.5 cm and 7.5 to 15 cm depths ranged from 12.3 to 20.7 mg kg−1 (figure 3b). Concentrations of 15 to 22 mg P kg−1 are considered high in Colorado soils, wherein yield responses to additional P are unlikely (Davis et al. 2009). However, in the 15 to 30 cm depth, STP in the control and urea treatments was <3 mg kg−1, well below sufficiency levels. These results indicate that despite multiple broadcast P applications, STP was below sufficiency in much of the rooting zone (i.e., depths below 15 cm) of the urea and control treatments due to low P mobility (Eghball et al. 1990). Manure application increased STP in all soil depths, where concentrations were ~500% higher than the control and urea treatments. Soil test P concentrations in the manure treatment were as high as 93.9 mg kg−1 in the surface 0 to 7.5 cm. Soil P levels in the 15 to 30 cm depth of the manure treatment were adequate, averaging 13.8 mg kg−1, indicating either movement of dissolved P from surface depths or enhanced P solubility due to chemical reaction of P with manure compounds (Eghball et al. 1996; Eghball et al. 2003).
These findings agree with previous studies demonstrating STP buildup with manure application (Eghball and Power 1999b; Halvorson et al. 2016; Schlegel et al. 2017). Schlegel et al. (2017) reported a buildup of STP after three years of N-based manure application to an alkaline soil (i.e., Olsen P levels of >100 mg kg−1); however, after 10 application years there was no additional evidence suggesting additional STP buildup had occurred. Previously at this study site, Halvorson et al. (2016) reported that fall STP levels in the surface 0 to 15 cm increased with manure addition, with STP concentrations of >110 mg kg−1 after three years of application (i.e., 2012 to 2014). The fact that spring postmanure application STP concentrations in 2018 did not increase beyond those reported for 2014 is surprising. In 2016, no manure was applied, which may have drawn down STP. The lack of additional increases in STP over time likely indicates soil P sorption into unavailable forms (Eghball et al. 2005). The influence of applied P on STP is dependent on initial P concentrations, application rates, sorption processes, and soil P saturation (Hansen et al. 2002). A large proportion of manure-borne P is inorganic, resulting in high P availability after application; however, P availability is a function of soil P sorptivity (Hansen et al. 2002). It is likely that P sorption processes (i.e., P adsorption to mineral surfaces and P precipitation) are high in this system due to high clay, pH, and calcium carbonate (CaCO3) contents. Phosphorus adsorption increases with clay content; in calcareous soils, high concentrations of Ca2+ in the soil solution, coupled with high pH, promotes the precipitation of calcium phosphates (Ca3(PO4)2), depressing phosphate (PO43−) availability (Eghball 2002; Eghball et al. 2005). The adsorption and immobilization of P by CaCO3 is especially rapid in calcareous soils (Mengel and Kirkby 1982). Overapplication of P carries multiple environmental risks of P losses via runoff or leaching; however, the high clay, high pH, calcareous soils of this region, coupled with distance from large water bodies, may reduce the environmental risk compared to other agricultural regions.
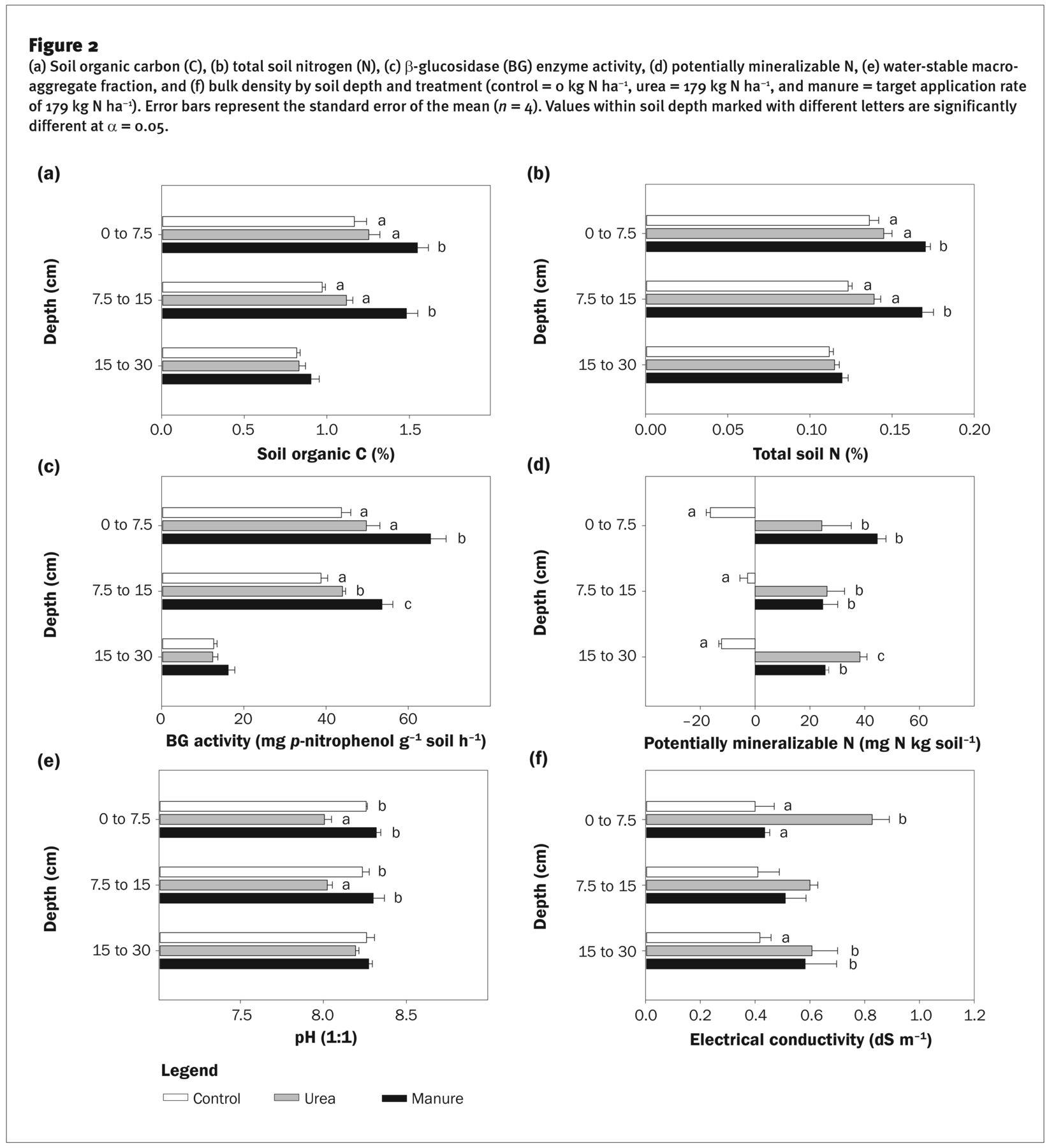
(a) Soil organic carbon (C), (b) total soil nitrogen (N), (c) β-glucosidase (BG) enzyme activity, (d) potentially mineralizable N, (e) water-stable macroaggregate fraction, and (f) bulk density by soil depth and treatment (control = 0 kg N ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 4). Values within soil depth marked with different letters are significantly different at α = 0.05.

(a) Soil nitrate nitrogen (NO3-N), (b) Olsen phosphorus, and (c) diethylenetriaminepentaacetic (DTPA) potassium by soil depth and treatment (control = 0 kg N ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 4). Values within soil depth marked with different letters are significantly different at α = 0.05.
Soil K concentrations were higher in the manure treatment than in the control and urea treatments in all depths (figure 3c). Manure contains large amounts of K, and over the course of six years, ~2,500 kg K ha−1 were applied via manure (table 1). Potassium contained in manure is estimated to be 100% plant available, and increases in soil K with manure application are frequently reported (Eghball et al. 2002; Lentz and Ippolito 2012; Schlegel et al. 2017; Vitosh et al. 1973). However, most Colorado soils are inherently high in extractable K, and very few growth responses to K fertilization have been reported.
Soil Micronutrients. Available soil Cu (Cuavail) ranged from 0.91 to 1.90 mg kg−1 between treatments (figure 4a), well above the critical threshold of 0.2 mg kg−1 identified for maize (Lindsay and Norvell 1978). Treatment differences were evident only in the 7.5 to 15 cm depth, where Cuavail in the manure treatment was higher than the urea treatment, but not different from the control (p = 0.07). Estimated total Cu applied annually in manure ranged from 1.2 to 2.5 kg ha−1, but less than half of total Cu was estimated to be available (table 1). Most Cuavail in calcareous soils is present as organic complexes, with soluble OM controlling the amount of complexed Cu in soil solution (Hodgson et al. 1966; McLaren and Crawford 1973). Manure application likely synergistically increases Cuavail via increased Cu inputs, as well as promotion of Cu complexing via increased OM. Others have also found increases in total and available Cu after manure application (Kornegay et al. 1976; Lentz and Ippolito 2012; Schlegel et al. 2017).
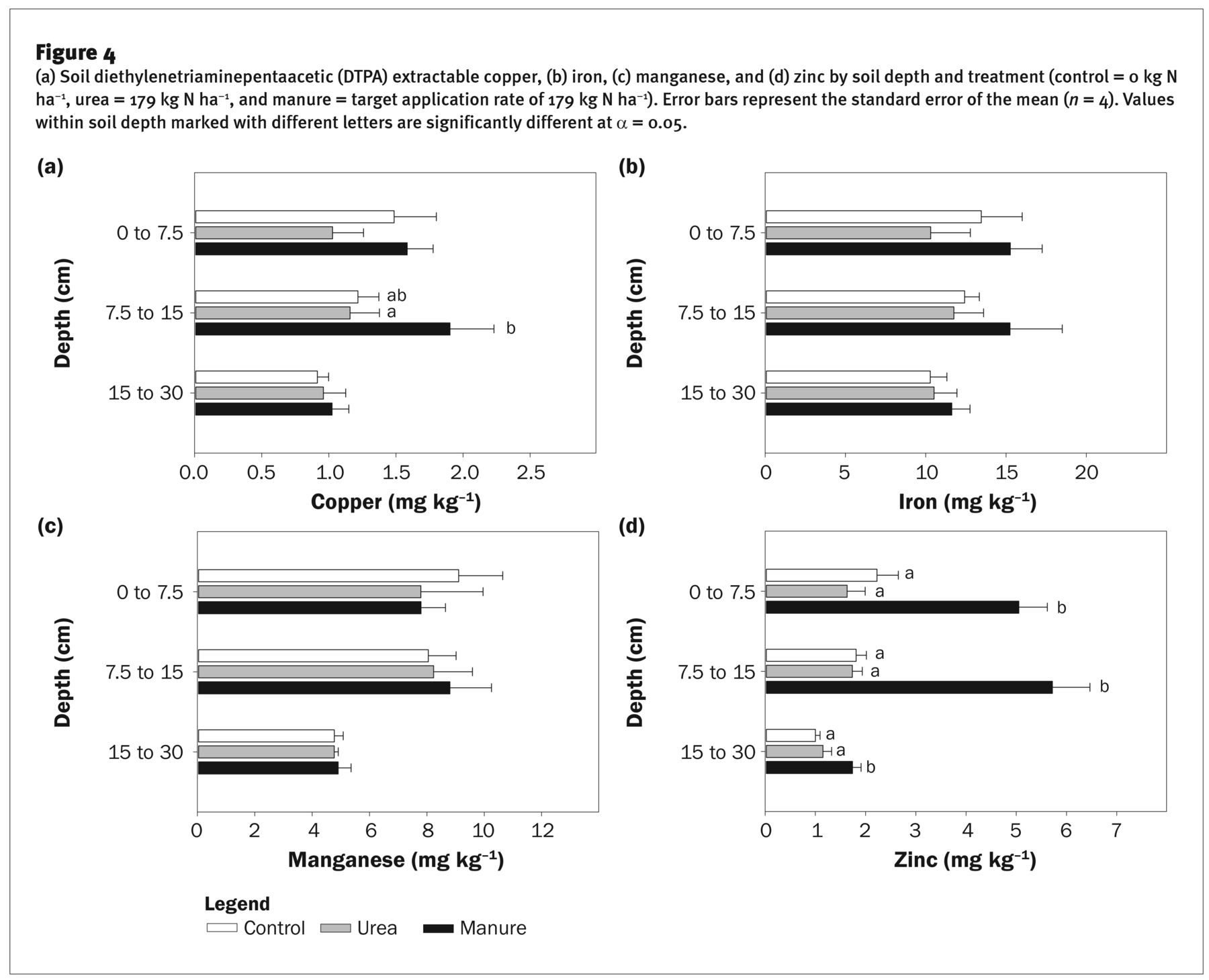
(a) Soil diethylenetriaminepentaacetic (DTPA) extractable copper, (b) iron, (c) manganese, and (d) zinc by soil depth and treatment (control = 0 kg N ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 4). Values within soil depth marked with different letters are significantly different at α = 0.05.
Available soil Fe (Feavail) ranged from 10.2 to 15.3 mg kg−1 (figure 4d). Iron availability in alkaline soils is sometimes limiting, yet Feavail levels were above sufficiency levels of 4.5 mg kg−1 (Lindsay and Norvell 1978). Although manure Fe content is infrequently reported, others have reported manure Fe of ≈0.5% (Lentz and Ippolito 2012), and in our analysis Fe concentrations ranged from ≈0.5% to 1% (table 2). Despite the annual addition of 225 to 550 kg ha−1 of total Fe with manure, there were no treatment differences in Feavail after six years of manure application (figure 4b). These results agree with other studies that have similarly reported no changes in Feavail after manure application (Lentz and Ippolito 2012; Schlegel et al. 2017). Most soils have a high abundance of total Fe, but at high soil pH, Fe is rapidly precipitated as Fe oxide and has very limited solubility (Lindsay and Schwab 1982), which likely explains the lack of treatment differences.
Available soil Mn (Mnavail) ranged from 4.8 to 9.1 mg kg−1 (figure 4c), well above the suggested sufficiency thresholds of 1.2 mg kg−1 (Lindsay and Norvell 1978). Despite the addition of an estimated 50 kg Mn ha−1 in manure over six years (table 1), there were no treatment differences in Mnavail. Other studies have also failed to find consistent increases in Mnavail with manure application (Lentz and Ippolito 2012; Schlegel et al. 2017). Lentz and Ippolito (2012) reported a 1.4-fold increase in Mnavail after a one-time manure application in the first growing season but could detect no differences relative to the control in the second season. Although increases in OM can increase solution and exchangeable Mn via chelation, adsorption and retention of Mn increases with increasing pH, clay, and SOM content, and Mnavail in the manure was likely adsorbed upon incorporation (Khattak and Page 2017; Mandal and Mitra 1982).
Available soil Zn (Znavail) ranged from 1 to 2 mg kg−1 in the check and urea treatments to as high as 5.7 mg kg−1 in the manure treatment (figure 4d). Levels of 1 to 1.5 mg kg−1 are considered marginal for maize, with >1.5 mg kg−1 considered adequate (Davis et al. 2009). An estimated 35 kg ha−1 of Zn was applied over the course of six years with manure, with ~50% estimated to be plant available (table 1). Zinc availability increased by 130% to 230% in the 0 to 7.5 cm and 7.5 to 15 cm depths with manure compared to the control or urea treatments. These results are consistent with other studies that have reported increases in Znavail with manure application (Schlegel et al. 2017). In addition, Znavail increased in the 15 to 30 cm depth, whereas the tillage operations in this system incorporate the manure only to 0 to 15 cm. These results suggest that Zn is leaching or moving in the soil profile when organic matter (manure) is added into the surface soil. Similar responses suggestive of Zn movement were found by Miner et al. (2018), who reported that Znavail increased in surface and subsurface soil depths after conversion to a no-till system increased surface soil C. Other authors have reported Zn movement in the soil profile with manure application (Asada et al. 2010; Jalali and Khanboluki 2007). Znavail is frequently low or limiting in alkaline soils—an estimated 50% of soils cultivated globally for cereal production are low in Znavail (Cakmak 2002), underscoring that increases in Znavail with manure would be highly beneficial in many cropping systems.
Soil Management Assessment Framework Soil Quality Indices. Physical, chemical, biological, nutrient, and overall SH indicator scores for the 0 to 7.5 cm and 7.5 to 15 cm depths are presented in figure 5. No differences were found in the 15 to 30 cm depth for SMAF indicators, except for in EC and PMN, so SMAF data are not presented for this depth.
The physical SH index scores in the 0 to 7.5 cm and 7.5 to 15 cm depths were ≥0.8 and did not differ between treatments. The physical SH index includes BD and aggregation, which did not change with treatment. The chemical SH index, which includes EC and pH, was greater in the urea treatment in both depths compared to the control or manure treatments, mainly due to lower soil pH. The three treatments separated with respect to biological SH due to the higher SOC and BG activity in the manure treatment and the higher mineralized N in the manure and urea treatments (figure 2). Despite the high STP levels in the manure treatment, the nutrient SH index was still ~0.8 and did not separate out from the urea and control treatments. The overall SH index for urea and manure treatments was higher than the control, but there were no differences between the urea and manure treatments, indicating no additional increases in overall SH despite the large inputs of OM and nutrients via manure.
The SMAF index rankings support the conclusion that adding manure and N fertilizer improves SH in this system. The SMAF nutrient SH index penalized the manure treatment due to high STP; however, the index did not account for the fact that although the STP was elevated with manure application, in this system, runoff or wind erosion were minimal, so this was an environmentally safe manure application. The SMAF index should include a separate category for environmental risk, so that cases with high nutrient levels that have low environmental risk (minimal/zero wind erosion and surface runoff) are more accurately represented (Delgado et al. 2008; Sharpley et al. 2003). In addition, the current tool considers only P and K in the nutrient SH index, so the benefits of the manure treatment to soil nutrient status (i.e., greater available Zn, a key nutrient for maize productivity) were not accounted for. Developing scoring function for additional macro- and micronutrients would improve the nutrient SH index and aid in the assessment of how management practices influence the availability of nutrients that can contribute to higher yields and improved crop quality.

Physical, chemical, biological, nutrient, and overall soil quality indices in the surface (a) 0 to 7.5 cm and (b) 7.5 to 15 cm by treatment (control = 0 kg N ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). The number of individual indicators used in each category is delineated in brackets. Error bars represent the standard error of the mean (n = 4). Values within soil depth and category marked with different letters are significantly different at α = 0.05.
Crop Productivity. Productivity was higher in the systems with higher overall SH (i.e., manure and urea treatments). Stover and cob yields were similar in 2017 and 2018. Averaged across years, the urea and manure treatments had comparable treatment responses, with both fertilizer treatments exceeding the control (figure 6). Grain yields were 10% higher in the urea and manure treatments in 2017 than in 2018 (p < 0.05). Grain yields did not differ between the urea and manure treatments and averaged 11,450 kg ha−1 over growing years, nearly 40% higher than the control, which averaged only 6,740 kg ha−1. These grain yields were similar to those reported by Halvorson et al. (2016) for the first three years of the study (2012 to 2014).
While the effect of manure versus inorganic N addition on yields is dependent on factors such as site, management, and manure quality and amount, grain yields in manure-based N treatments are often equal to or greater than inorganic N treatment yields (Eghball and Power 1999a; Jokela 1992; Ma et al. 1999). We found no differences between inorganic and organic N sources on stover, cob, or grain yields. These results align with previous work in this system (Halvorson et al. 2016) and indicate that manure is as effective as inorganic N in supporting high yields in this irrigated, tilled system.

Average stover, cob, and grain yields in response to inorganic nitrogen (N) (179 kg N ha−1) and manure treatment (estimated available N of 179 kg N ha−1), compared to a control treatment (0 kg N ha−1). Data are the mean ±1 SE averaged over two growing seasons (n = 8). Different lowercase letters within a plant fraction indicate a significant treatment difference at α = 0.05.
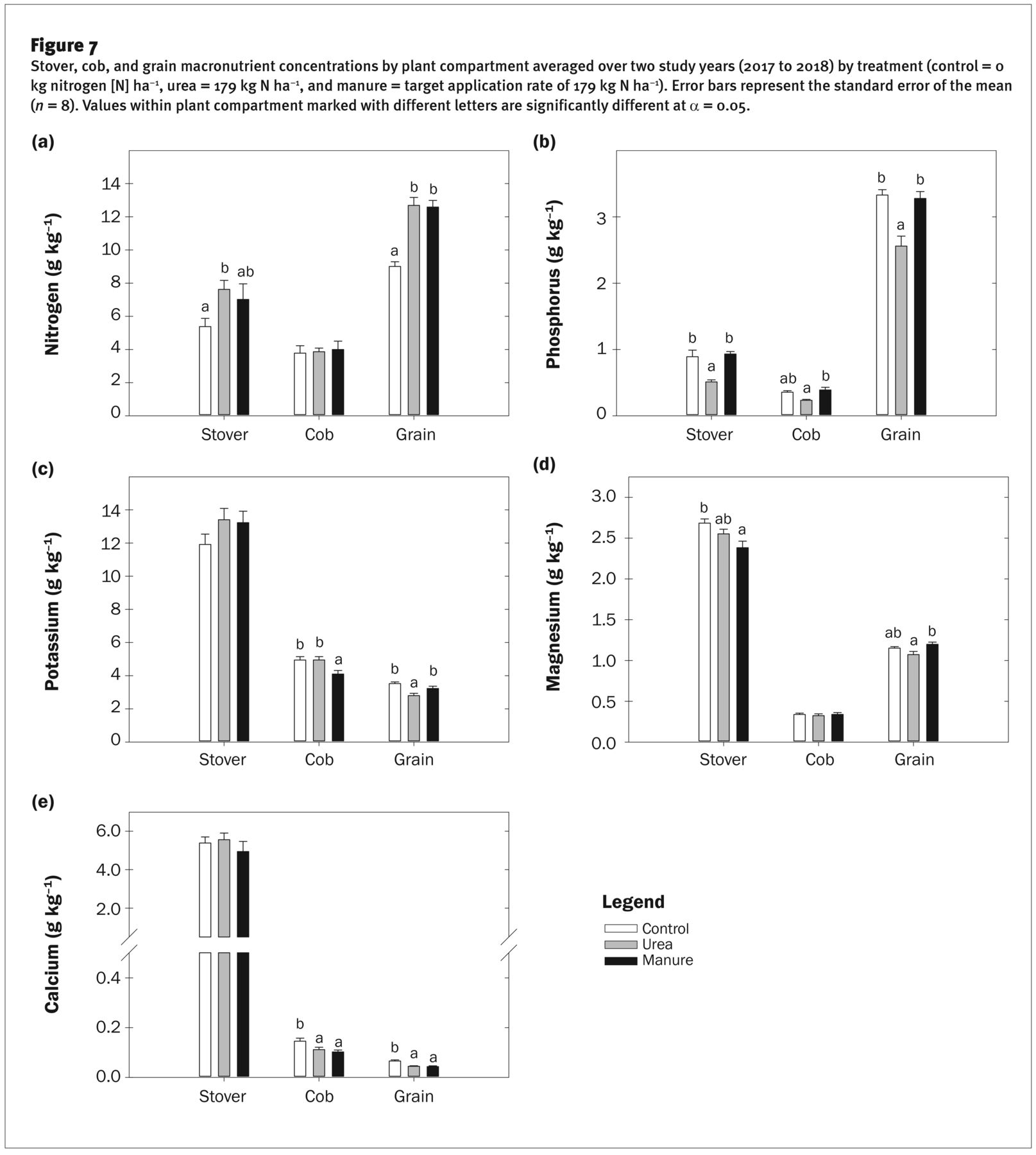
Crop Quality: Nutrient Concentrations and Uptake. Stover [N] was higher in 2017 than in 2018 for all treatments (p ≤ 0.001), averaging 8.1 g kg−1 in 2017 versus 5.3 g kg−1 in 2018. Stover [N] in the urea treatment was higher than the control, whereas stover [N] in the manure treatment did not differ from the control (p = 0.13) (figure 7a). Cob [N] was higher in 2017 than in 2018 (i.e., 4.8 versus 3 g kg−1), but it was not altered by treatment (figure 7a), consistent with the findings of Halvorson and Johnson (2009). Grain [N] was similar in both 2017 and 2018, but it was 40% higher in the urea and manure treatments as compared to the control, reaching an upper threshold of ~13 g kg−1 (figure 7a). This maximum grain [N] is similar to values reported by others for modern maize genotypes (Blesh and Drinkwater 2013; Miner et al. 2018; Woli et al. 2018). The similar stover and grain [N] between the urea and manure treatment indicate that, despite low spring soil NO3-N in the manure treatment, seasonal mineralization rates were sufficient to meet crop N demands (i.e., figure 2d). The increases in stover and grain [N] in the manure and urea treatments, coupled with higher yields, markedly increased total N uptake over the control (i.e., by >100 kg N ha−1) (table 3).
Stover [P] was similar in both growing years, but it was nearly twice as high in the manure and control treatments (>0.90 g kg−1) as in the urea treatment (0.51 g kg−1) (figure 7b). Stover [P] in the control and manure treatments was slightly lower than concentrations reported by others (Ciampitti et al. 2013; Mallarino et al. 2011). The lack of increase in stover [P] in the manure treatment despite high STP levels suggests an upper limit of stover [P] accumulation, with little or no luxury uptake, which concurs with the results of Mallarino (1996) who found clear upper limits of P uptake in young plants and leaves “only slightly higher than values needed to produce maximum economic yields.” Cob [P] was similar between growing years but was higher in the manure treatment than in the urea treatment (figure 7b). Grain [P] was 10% and 25% lower in 2017 than in 2018 in the manure and urea treatment, respectively, with no interannual differences in the control. Across growing years, grain [P] in the control and manure treatments averaged 3.32 and 3.28 g kg−1, respectively, versus only 2.56 g kg−1 in the urea treatment.
The bulk of plant P accumulates in the grain (Abendroth et al. 2011), and hence high grain [P] in the manure treatment coupled with high yields resulted in higher total P uptake than in the check or urea treatments (table 3). Indeed, we found that <2% of total plant P was in the cob, 12% to 18% was in the stalk, and 80% to 87% of total plant P was contained in the grain. Total P uptake averaged 44.5 kg ha−1 in the manure treatment, versus only 28.6 and 31.9 kg ha−1 in the check and urea treatments, respectively.

Stover, cob, and grain macronutrient concentrations by plant compartment averaged over two study years (2017 to 2018) by treatment (control = 0 kg nitrogen [N] ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 8). Values within plant compartment marked with different letters are significantly different at α = 0.05.
Our results suggest that manure application in this system supports luxury grain P uptake (i.e., increases in [P] that do not increase yield). Conversely, the high yields in the urea treatment resulted in a dilution of grain [P], which is an interesting result as multiple studies have reported that grain [P] is not readily subject to yield dilutions. For example, Mallarino et al. (2011) found no relationship between grain [P] and yield across dozens of years, sites, and treatments, with an average grain [P] of 3 g kg−1. Ciampitti et al. (2013) reported that grain [P] was stable across multiple N treatments despite yield variation, averaging 3.6 g kg−1. Feil et al. (2005) found no effect of N fertilization on grain [P] in two growing years, with an average grain [P] of 3.23 g kg−1. Eghball et al. (2003) reported large variation in grain [P] between hybrids (i.e., 2.12 to 3.28 g kg−1) but no impact of N fertilization or yield on grain [P]. However, STP levels in these studies were likely well above sufficiency levels due to soil pH levels and P additions.
Macronutrient uptake (kg ha−1) for maize (Zea mays L.) by plant fraction for the control treatment, inorganic nitrogen (N) treatment (179 kg N ha−1), and manure treatment (target seasonal N availability of 179 kg N ha−1) over the 2017 and 2018 growing years. Different lowercase letters within nutrient indicate a significant difference between treatments.
In our study, while STP levels in the control and urea treatments were above the levels wherein yield responses are likely, they were likely not high enough to promote luxury P uptake. For example, Barber (1979) examined changes in grain [P] with five P rates and found that while yield was maximized at ~11 kg P ha−1, grain [P] continued to increase with additional P fertilizer additions, indicating luxury P consumption when STP was high. Mallarino (1996) evaluated grain [P] in response to P fertilization and reported that while grain [P] was poorly related to yield, a relationship existed between grain [P] and surface and subsoil STP in unfertilized plots, with an increasing relative importance of subsoil P as the growing season advanced. In our study, manure application increased grain [P] either due to an abundance of STP in the 0 to 15 cm depth or due to sufficiency in the subsoil (Mallarino 1996; figure 3b). Interestingly, we found that grain P never rose above 3.42 mg kg−1 despite very high STP values coupled with irrigated conditions that likely maximized soil P diffusion. Previously published values for expected grain [P] ranged from 2 to 6 mg kg−1 (Walsh and Beaton 1973). However, our results, coupled with other published results noted above, suggest that the upper limit for grain [P] is in the range of 3.5 to 4 mg kg−1.
Understanding the conditions that promote luxury P uptake, as well as upper limits for grain [P], may be important for animal and human nutrition. Luxury P accumulation is desirable if the goal is to remove P in harvested products and thereby reduce P accumulation in soils (Eghball et al. 2003). A large proportion of the maize grown in the United States is used for animal feed. Phosphorus is an essential element in animal nutrition (Kincaid et al. 1981), and while requirements for key minerals such as P are affected by a various dietary and animal factors, mineral P supplements are incorporated into diets for many classes of livestock (e.g., swine, poultry, feedlot cattle, and dairy cows) (McDowell 1996). The majority of P in maize grain (i.e., 70% to 80%) occurs as mixed K and Mg salts of phytic acid, commonly known as phytate (Lott et al. 2000; Raboy 2001). Although phytate is a poor P source for monogastrics, there is now widespread use of phytase enzymes in swine and poultry feed, which increase the bioavailability of phytate. Ruminants such as cattle can release this bound P due to phytase-generating bacteria in the rumen. High stover and grain [P] hence reduce the amount of supplemental P that is needed (Kleinman et al. 2018). Maize is also processed for human consumption. While there is some evidence that in humans phytic acid can positively act as an anticancer agent and an antioxidant (Feil 2001), phytate also functions as an “antinutrient,” chelating and reducing the bioavailability of minerals such as Ca, Fe, and Zn, which can pose public health problems in populations at risk of Fe and Zn deficiencies (Raboy 2001). In human nutrition, increases in grain [P] can hence represent either positive or negative shifts in quality, depending on the target population (Lott et al. 2000).
Stover [K] was higher in 2018 than in 2017 (i.e., 14 versus 11.5 g kg−1; p ≤ 0.05). Stover [K] was not influenced by treatment, despite the higher soil K in the manure treatment (figure 7c). Stover [K] averaged 12.9 g kg−1 over treatments and years, similar to the value reported by Mallarino et al. (2011). Cob [K] was slightly lower in the manure treatment (4.1 g kg−1) than in the control and urea treatments (4.9 g kg−1). Grain [K] was lower in 2018 than in 2017 (i.e., 3.4 versus 2.9 g kg−1, respectively; p ≤ 0.05). Grain [K] averaged 2.8 g kg−1 in the urea treatment versus 3.2 and 3.5 g kg−1 in the manure and check treatments (p < 0.05). Grain [K] typically ranges from 2 to 5 g kg−1 (Bruns and Ebelhar 2006; Ciampitti et al. 2013; Feil et al. 2005). While other studies have found no dilution of grain [K] with yield (Ciampitti et al. 2013; Mallarino et al. 2011), our results suggest both a dilution of grain [K] with urea fertilization as well as a synergistic effect of manure on grain [K] (i.e., 15% higher grain [K] despite no yield differences). Although we did not measure phytate directly, phytate is a mixed cation salt, and the negatively charged sites of phytic acid typically bind with K and Mg (Marschner 1995). Hence, increases in grain [P] with manure would be expected to synergistically increase grain [K] and [Mg]. Total K uptake ranged from 100.7 kg ha−1 in the control treatment to 151.1 kg ha−1 in the manure treatment (table 3).
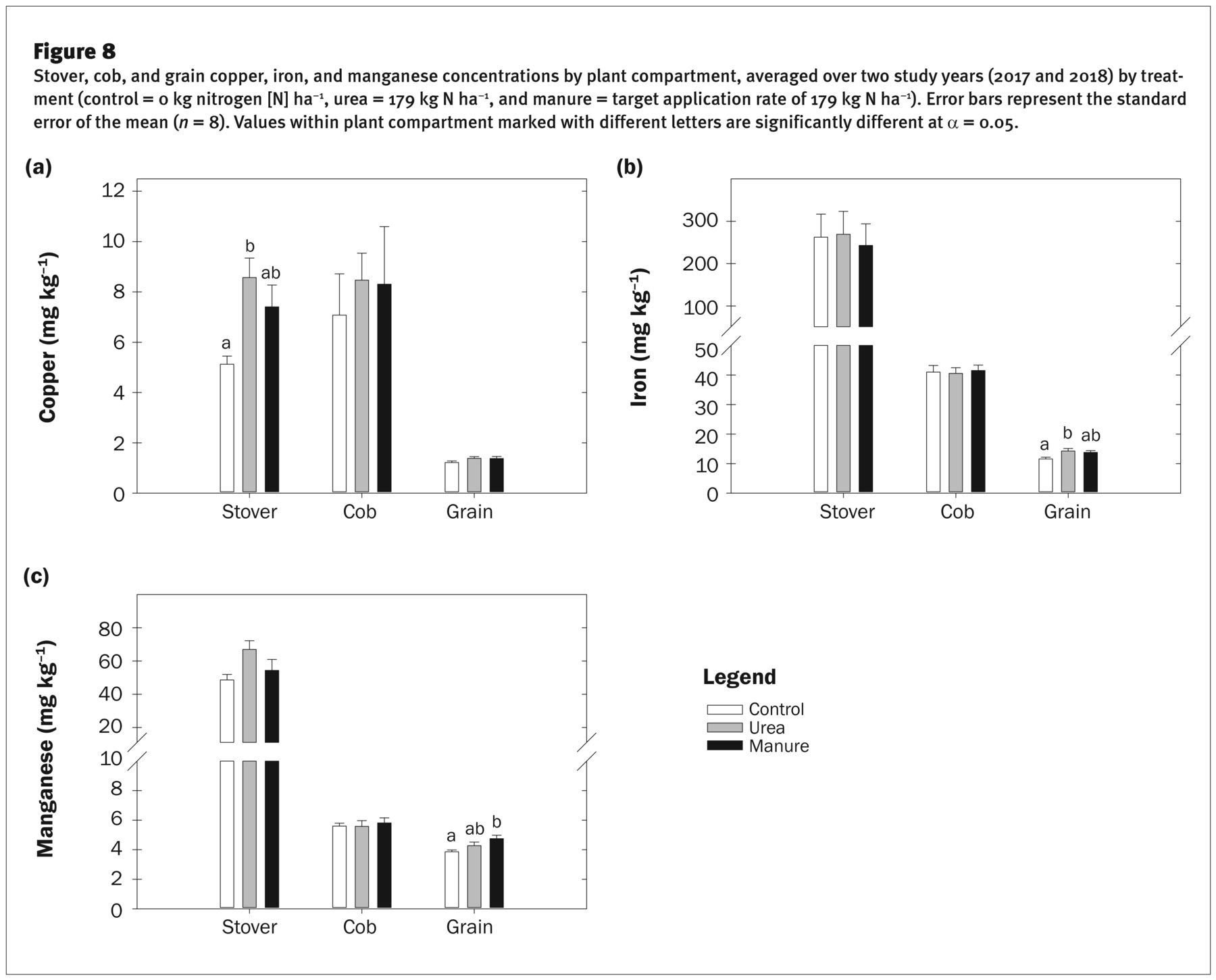
Stover, cob, and grain copper, iron, and manganese concentrations by plant compartment, averaged over two study years (2017 and 2018) by treatment (control = 0 kg nitrogen [N] ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 8). Values within plant compartment marked with different letters are significantly different at α = 0.05.
Stover [Mg] did not vary between growing years and ranged from 2.38 to 2.68 g kg−1 (figure 7d). These stover Mg concentrations are 0.5 to 1 g kg−1 higher than those reported by others (Bruns and Ebelhar 2006; Ciampitti and Vyn 2013; Feil et al. 2005). Stover [Mg] was lower in the manure treatment than in the control (p = 0.02). Cob [Mg] did not vary between treatments and averaged only 0.32 g kg−1. Grain [Mg] ranged from 1.07 to 1.2 g kg−1 and was 10% lower in the urea treatment than in the manure treatment. Feil et al. (2005) reported similar grain [Mg] (i.e., 1.12 g kg−1), but found no change in grain [Mg] with N fertility or yield, whereas Ciampitti and Vyn (2013) noted grain [Mg] ranging from 1.35 to 1.71 g kg−1 and found increases in grain [Mg] in response to N. We found that grain [Mg] was highest in the manure treatment, likely due to the same mechanisms posited for increases in grain [K]. Total uptake ranged from 24.3 to 33.4 kg ha−1 and was highest in the urea and manure treatments (table 3).
Stover [Ca] was ~25% higher in 2017 than in 2018 (i.e., 5.85 g kg−1 versus 4.74 g kg−1; p = 0.04) but was not impacted by treatment (figure 7e). Some studies have reported increases in stover [Ca] with N fertilization (Ciampitti and Vyn 2013), but we did not observe this, likely due to high soil Ca concentrations. The stover [Ca] we measured was approximately twice as high as those reported by others (Bruns and Ebelhar 2006; Ciampitti and Vyn 2013). High levels of Ca in the external soil solution typically increase the Ca concentrations in plant leaves, whereas these effects are not necessarily seen in low-transpiring organs supplied via the phloem (i.e., grain) due to restriction of Ca transport in the phloem stream (Marschner 1995). Indeed, cob [Ca] was <3% of stover [Ca], averaging 0.10 g kg−1 in the manure and urea treatment versus 0.14 g kg−1 in the control. Grain [Ca] was <1% of stalk [Ca], averaging 0.04 g kg−1 in the urea and manure treatments versus 0.06 g kg−1 in the control. A dilution in grain [Ca] with N fertility was also reported by Feil et al. (2005), whereas other studies have not found this effect (Ciampitti and Vyn 2013). Total uptake ranged from 32.5 to 45.9 kg ha−1 and did not differ between treatments (table 3).
Stover [Cu] ranged from 5.1 to 8.6 mg kg−1 (figure 8a), similar to the levels reported by others (Berenguer et al. 2008; Bruns and Ebelhar 2006; Losak et al. 2011; Miner et al. 2018). Stover [Cu] was higher in the urea treatment than in the control (figure 8a). However, despite the increase in Cuavail with manure, stover [Cu] in the manure treatment did not differ from the control or urea treatment. Other studies have also reported no differences in stover [Cu] between manure and inorganic N treatments (Berenguer et al. 2008; McIntosh and Varney 1972). Interestingly, multiple studies have found that when soil Cu concentrations are elevated, maize plants can accumulate very high levels of Cu in the roots (i.e., 60 to 600 mg kg−1) and yet tightly restrict translocation to shoots to keep aerial and grain concentrations low (Brun et al. 2001; Jarausch-Wehrheim et al. 1996; Mantovi et al. 2003). Translocation of Cu from the roots to the aerial organs is thought to be controlled by N metabolism, with Cu moving in the xylem complexed with organic-N ligands or chelated to amino acids (Marschner 1995). Hence, providing soil Cu levels are not deficient, additional increases in soil Cu due to manure application should not increase stover [Cu]; rather stover [Cu] should correlate with N uptake and movement. Indeed, the correlation between stover [Cu] and [N] was r > 0.90 in both growing years. These results are consistent with other studies that have found a synergistic relationship between stover [N] and [Cu] (Bruns and Ebelhar 2006; Ciampitti and Vyn 2013; Miner et al. 2018).
The higher stover [Cu] in the manure and urea treatments did not result in higher cob or grain [Cu] than in the control (figure 8a). Grain [Cu] measured <1.4 mg kg−1. While Cu mobility from the vegetative tissues to the grain via the phloem is not well understood (Grusak et al. 1999), we found no evidence of increased Cu remobilization with higher source Cu in maize, nor did grain [Cu] parallel increases in grain [N]. The percentage of Cu in the grain was <18% of total plant Cu in all three treatments. Our results agree with other studies that have reported increases in stover [Cu] with increased N fertility, with no corresponding changes in grain [Cu] (Bruns and Ebelhar 2006; Miner et al. 2018). Maximum total Cu uptake was ≈0.1 kg ha−1 (table 4).
Stover [Fe] was not influenced by treatment (figure 8b) but differed between growing years (p = <0.001). Stover [Fe] was 225% higher in 2017 than in 2018, averaging 394.5 mg kg−1 in 2017 versus only 121.7 mg kg−1 in 2018. While the differences in genotypes between growing years make it impossible to attribute differences entirely to growing year, Miner et al. (2018) also reported an 80% difference in stover [Fe] between two growing years where the same maize hybrid was grown, suggesting that factors such as soil and growth conditions (i.e., temperature and moisture) may contribute to large interannual differences in stover [Fe] (Fageria et al. 2002). Cob [Fe] was lower in 2017 than in 2018 (i.e., 38.4 ± 3.2 mg kg−1 versus 43.6 ± 6 mg kg−1; p = 0.048) but was not influenced by treatment. Despite the higher stover [Fe] in 2017, grain [Fe] was lower in 2017 than in 2018 (i.e., 12.6 mg kg−1 versus 13.9 mg kg−1; p = 0.03). Grain [Fe] was influenced by treatment, with higher grain [Fe] in the urea versus the check treatment (p = 0.04), whereas differences between the check and manure treatments were not significant (p = 0.09). There was a strong correlation between grain [Fe] and grain [N] in both growing years (r = 0.70 and r = 0.83), which concurs with the grain [N]−[Fe] synergism reported by Miner et al. (2018). Interestingly, while the processes of N remobilization to the grain are largely understood (Hay and Porter 2006), comparatively little is known about the processes of Fe movement or remobilization to the grain (Grusak and DellaPenna 1999). This synergism could perhaps be due to factors that are improved with N nutrition (i.e., root proliferation and/or longevity, canopy photosynthetic capacity and longevity), simultaneously supporting Fe uptake and movement (Marschner 1995).
Micronutrient uptake (g ha−1) for maize (Zea mays L.) by plant fraction for the control treatment (0 kg N ha−1), inorganic nitrogen (N) treatment (179 kg N ha−1), and manure treatment (target seasonal N availability of 179 kg N ha−1) over the 2017 and 2018 growing years. Different lowercase letters within nutrient indicate a significant difference between treatments.
These results imply that grain [Fe] is not controlled solely by [Fe] in vegetative source tissues, and stover [Fe] may not reflect Fe available for transport to the grain. Other studies have demonstrated that total leaf Fe content is of limited value as an indicator of Fe nutritional status—only a fraction of Fe is thought to be physiologically active, with a large amount of Fe accumulated in unavailable forms (Abadía 1992; Marschner 1995). In addition, phloem loading and transport of Fe from the stover to the grain is limited by the need for a specific chelator (Grusak et al. 1999), which may also limit final grain [Fe]. Interestingly, Fe partitioning and movement between vegetative and reproductive organs varies widely between crop species. For example, Fe deposition to the seed in pea (Pisum sativum L.) was found to represent 75% of total shoot Fe, whereas in rice (Oryza sativa L.) only 4% of shoot Fe was partitioned to the grain (Grusak and DellaPenna 1999). In 2017, we found that only 3% to 5% of stover Fe was partitioned to the grain in 2017, versus 12% to 17% in 2018. Despite high stover Fe contents, only a small amount of Fe was remobilized to the grain in maize. Total Fe uptake ranged from 1,681 to 2,445 g ha−1 (table 3).
Stover [Mn] was ~25% higher in 2017 than in 2018 (p = 0.04), whereas cob and grain [Mn] did not differ between growing years. Averaged over year, stover [Mn] ranged from 48.6 to 67 mg kg−1 between treatments (figure 8c), well above published sufficiency levels of 10 to 25 mg kg−1 (Mengel and Kirkby 1982) and similar to concentrations reported elsewhere (Ciampitti and Vyn 2013; Miner et al. 2018). Higher stover [Mn] in 2017 likely reflects seasonal differences in environmental variables that influence Mn and water availability to the roots and transpiration, as Mn is easily mobilized from the root to the shoot in the xylem (Kochian 1991). Treatment impacts on plant [Mn] were only evident in the grain, where grain [Mn] in the manure treatment averaged 4.8 mg kg−1, 23% higher than the control (3.9 mg kg−1) but not different from the urea treatment. Maximum total Mn annual uptake was ~0.6 kg Mn ha−1 (table 3).
Several studies have reported no changes in grain [Mn] with higher N fertility (Brun et al. 2001; Losak et al. 2011; Miner et al. 2018), while others have found increased grain [Mn] with N fertilization (Ciampitti and Vyn 2013; Feil et al. 2005). We found a detectable synergistic treatment impact of manure on grain [Mn].
Despite the increase in Znavail with manure application, there were no treatment differences in stover [Zn] in either growing year (figure 9). Stover [Zn] averaged 37.8 mg kg−1, well within the whole plant [Zn] sufficiency range of 20 to 50 mg kg−1 (Mengel and Kirkby 1982) and similar to other published values (Bruns and Ebelhar 2006; Ciampitti and Vyn 2013; Miner et al. 2018). The interactions between Zn and N nutrition in the published literature are not consistent; some studies have found declines in stover [Zn] with increasing N fertility (Ciampitti and Vyn 2013; Miner et al. 2018), while others have reported no changes in stover [Zn] with N (Bruns and Ebelhar 2006; Losak et al. 2011). Cob [Zn] was impacted by treatment (p < 0.05) but not growing year.
There was an interaction between year and treatment on grain [Zn] (p = 0.03) (figure 9). Grain [Zn] ranged from 17.4 to 23.2 mg kg−1 between treatments and growing years, similar to other published values (Berenguer et al. 2008; Bruns and Ebelhar 2006; Losak et al. 2011; Miner et al. 2018). Grain [Zn] levels were approximately 2 to 5 mg kg−1 higher in 2017 than in 2018 in the control and manure treatments, whereas grain [Zn] did not vary between years in the urea treatment. In 2017, grain [Zn] declined in both the urea and manure treatments compared to the control, whereas in 2018 there were no treatment differences in grain [Zn]. These interannual differences in N treatment impacts on grain [Zn] suggest either curtailment of Zn uptake and/or remobilization during the grain filling period in 2017, or prolonged remobilization of carbohydrates from leaves and stalk to the grain that was not matched by Zn uptake and/or remobilization. Reported impacts of N on grain [Zn] are disparate, with multiple studies reporting no change in grain [Zn] with N fertility (Bruns and Ebelhar 2006; Losak et al. 2011; Riedell et al. 2009), whereas others have found reductions in grain [Zn] with increasing N (Bruns and Ebelhar 2006; Feil et al. 2005; Li et al. 2007; Miner et al. 2018; Riedell et al. 2009). Zinc concentrations in multiple cereal grains have been shown to increase in response to Zn additions, even when yield responses are not evident (Marschner 1995; Rengel et al. 1999), which suggests that the variability in observed responses is likely due to differences in available soil Zn between studies. Total Zn uptake ranged from 0.40 to 0.53 kg ha−1 between treatments (table 3).

Stover, cob, and grain zinc concentrations by plant compartment for (a) 2017 and (b) 2018 by treatment (control = 0 kg nitrogen [N] ha−1, urea = 179 kg N ha−1, and manure = target application rate of 179 kg N ha−1). Error bars represent the standard error of the mean (n = 4). Values within plant compartment marked with different letters are significantly different at α = 0.05.
Summary and Conclusions
We examined the connections between SH, crop productivity, and crop quality in a continuous maize system with treatments varying in input type and amount. The overall SH index was higher in the manure and urea treatments than in the control, but no differences in this index between the organic and inorganic N sources were found despite the large exogenous sources of C, N, and nutrients applied via manure. The biological SH index was higher for the manure-treated soils than the urea and control treatments, reflecting the increases in SOC and enzyme activities with manure addition. The SMAF nutrient index penalized the manure treatment due to high STP; however, in this study, runoff or wind erosion were minimal, so this should be considered an environmentally safe manure application, even if high STP were measured. In addition, the current SMAF tool considers only P and K in terms of soil nutrients—the nutrient benefits of manure application (i.e., higher TSN, increases in available soil micronutrients) were not accounted for, suggesting that the SMAF nutrient SH index should be modified to account for the impacts of management practices on nutrient availability.
Both the inorganic and organic N amendments improved SH and crop productivity over the control treatment. Despite widely differing nutrient inputs and soil fertility levels, there were no yield differences between urea and manure treatments, indicating that manure was as effective as inorganic N in supporting yields in this system. However, there were notable differences in crop quality between treatments. Stover [N] was 30% to 50% higher in the urea and manure treatments than in the control, and stover [P] was 80% higher in the manure treatment than in the urea treatment. These differences in stover composition will impact the quality of silage in silage production systems. Grain [N] was 40% higher in the urea and manure treatments than in the control. Grain P, K, and Mg, important elements in livestock nutrition, were also higher in the manure treatment than in the urea treatment, indicating improvements in crop nutritional quality and reductions in supplement costs for producers. In addition, the Zn dilution effect observed in the urea treatment was slightly mitigated in the manure treatment, likely due to increases in available soil Zn. Hence, although the overall SH was the same between the urea and manure treatments, there were additional increases in crop quality in the manure treatment that were not captured by the SH index. While this work examines the interrelationships between management, productivity, and crop quality for maize, understanding these relationships for other staple crops is a critical component of combating food system nutrient deficiencies at the field level. Our results suggest that management practices that support soil health and nutrient availability also support the productivity and nutritional quality of maize, which could have positive impacts on animal and human nutrition.
- Received June 11, 2019.
- Revision received September 30, 2019.
- Accepted November 5, 2019.
- © 2020 by the Soil and Water Conservation Society





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}